Induction of Cytomegalovirus pp65-specific CD8+T Cells in vivo Following Administration of Autologous Tumorlysate-pulsed Dendritic Cells, Coplused with Zoledronate and α-Galactosylceramide in a Patient with Glioblastoma multiforme
Motoki Yagi1, Hiroshi Terunuma1,2,3, Yuuta Eiraku1, Xuewen Deng1, Mie Nieda1
2.Department of Immunotherapy, Tokyo Clinic, 1F Shin-Otemachi, 2-2-1 Otemachi, Chiyoda-ku, Tokyo 100-0004, Japan.
3.Department of Immunotherapy, Southern Tohoku General Hospital, Fukushima 963-8052, Japan.
Citation : Yagi M, Terunuma H, Eiraku Y, Deng X, Nieda M. Induction of Cytomegalovirus pp65-specific CD8+T Cells in vivo Following Administration of Autologous Tumorlysate-pulsed Dendritic Cells, Coplused with Zoledronate and Galactosylceramide in a Patient with Glioblastoma multiforme . Clin Res Immunol 2018;1(1):1-5.
Abstract
Vγ9γδT cells, innate immune cells activated by zoledronate (Z), and Va24 natural killer T cells, and innate/adaptive immune cells activated by a-galactosylceramide (G) can link innate and adaptive immunities through cross talk with interferon (IFN) a-derived dendritic cells (DCs) (IFN-DCs) from patients with GBM in a way that can amplify the activation and proliferation of cytomegalovirus (CMV) pp65-specific CD8+T cells and that can reduce proliferation of CD4+CD25+FoxP3+regulatory T cells (Treg cells) in vitro. In this study, we demonstrate for a patient with glioblastoma multiforme (GBM) that multiple administrations of IFN-DCs pulsed with autologous tumor lysates and Z plus G to the patient result in the enhancement of CMV pp65-specific CD8+T cells and in the reduction of the Treg cells in vivo. This leads to the higher induction of CMV pp65-specific CD8+T cells by IFN-DCs pulsed with CMV pp65 peptide and Z plus G derived from peripheral blood mononuclear cells (PBMCs) after DC therapy, compared with that by IFN-DCs pulsed with the peptide and Z plus G derived from PBMCs before DC therapy in vitro. Our results indicate that administration of IFN-DCs pulsed with autologous tumor lysates and Z plus G may lead to a good clinical outcome for patients with GBM.
α-galactosylceramide, cytomegalovirus pp65-specific CD8+T cells, cytomegalovirus pp65, glioblastoma multiforme, zoledronate
INTRODUCTION
Glioblastoma multiforme (GBM) is the most common and malignant primary brain tumor in adults. Despite advancing treatments including resection, radiotherapy, and chemotherapy, most patients survive < 1 year [1-3]. Recently, an association has been discovered between human cytomegalovirus (CMV) and malignant gliomas in vivo and an extremely robust human CMV-specific CD8+T cell response to the CMV pp65 immunodominant epitope was observed after one injection of autologous tumor lysate-pulsed dendritic cell (DC) [4]. This rapid expansion of an anti-CMV immune response was not associated with any systemic symptoms of viral illness possibly due to the selective tropism of human CMV for glial cells. It has been also reported that CMV pp65 is an optimal target in GBM as the tumor antigen as CMVpp65 have been detected in cancellous areas of histological sections but not in surrounding healthy tissues [5-7]. We have recently reported that Vγ9γδT cells, innate immune cells activated by zoledronate (Z), and Va24 natural killer T (NKT) cells, and innate/adaptive immune cells activated by a-galactosylceramide (G) can link innate and adaptive immunities through cross talk with interferon (IFN)-DCs from patients with GBM in a way that can amplify the activation and proliferation of CMV pp65-specific CD8+T cells in vitro and that can reduce the proliferation of
CD4+CD25+FoxP3+ regulatory T cells (Treg cells) in vitro [8].
Based on these observations in vitro together with a previous study using tumor lysates from a patient with GBM in vivo,[4] we have done with DC therapy for patient 1 with GBM using IFN-DCs pulsed with autologous tumor lysates and Z plus G. The results showed that multiple administrations of IFN-DCs pulsed with autologous tumor lysates and Z plus G induced enhancement of CMV pp65-specific CD8+T cells and reduction of the Treg cells in vivo. This resulted in the higher induction of CMV pp65-specific CD8+T cells by IFN-DCs pulsed with CMV pp65 peptide and Z plus G using peripheral blood (PB) mononuclear cells (PBMCs) after DC therapy, compared with that by IFN-DCs pulsed with the peptide and Z plus G using PBMCs before DC therapy in vitro.
MATERIALS AND METHODS
Surface phenotypes were determined using an Navios (Beckman Coulter, CA). The following monoclonal antibody (mAbs) was purchased from Beckman Coulter: anti-CD3, anti-CD4, anti-CD8, anti-Vγ9T cell receptor (TCR), anti-CD14, anti-CD25, anti-CD45, anti-CD56, anti-human leukocyte antigens (HLA) DR, anti-CD40, anti-CD80, anti-CD86, anti-CD11c, anti-CD36, mouse IgG1, mouse IgG2, and mouse IgG2b mAbs. Anti-HLA-Class 1 and anti-CCR7 mAbs were purchased from Becton Dickinson (San Jose, CA) and R and D system (Minneapolis, MN), respectively. Anti-TCR Va24 mAbs and anti-TCR VΒ11 mAbs were purchased from Beckman Coulter (France). Anti-FoxP3 mAb for intracellular staining was purchased from BD Biosciences (Japan). All mAbs were conjugated with fluorescein isothiocyanate (FITC), PE, APC, ECD, PC5, or PC7. Z was purchased from Novartis Pharmaceuticals (Switzerland) and G from Funakoshi Co., Ltd. (Japan).
PBMCs were isolated from HLA-A*0201 positive patient 1 with GBM. The subject provided their written informed consent to use their PBMCs for research purpose. For cell culture in this study, AIM-V medium (Invitrogen, Japan) containing 10% heat-inactivated fetal bovine serum for in vitro study or autologous plasma for in vivo study was used. PBMCs separated by density gradient centrifugation with Lymphoprep (Nycomed, Norway) were suspended in the medium and then incubated in a flask (Corning Incorporated, Japan) for 1h at 37°C. After removing non-adherent cells containing CD8+T cells, Vγ9γδT cells, and Va24NKT cells, monocytes obtained as adherent cells were cultured in the medium containing granulocyte macrophage colony-stimulating factor (GM-CSF) (1000 U/mL; Primmune Inc., Japan) and IFNa (1000 U/mL; INTRON, MSD K.K., Japan) for 3 days to obtain IFN-DCs.
Tumor lysates were prepared by multiple freeze and thaw cycle method described previously [9]. IFN-DCs were incubated with tumor lysates (100 µg/ml) overnight and then washed with PBS twice to remove excess tumor lysates to obtain tumor lysate-pulsed IFN-DCs.
To investigate MHC-restricted immune responses mediated by CD8+T cells before DC therapy and after DC therapy in vitro, HLA- A*0201 restricted, CMV pp65-9-mer synthetic peptides (NLVPMVATV) were used. Lymphocytes (2 X 106) as a non-adherent cell fraction were cultured with autologous IFN-DCs (2 X 105) under two different conditions. IFN-DCs were pulsed with the HLA-A*0201-restricted CMV pp65 peptide (5 µg/mL) for 2h in serum-free medium and then cultured with the lymphocytes possessing HLA-A*0201. In some experiments, Z (0.1 µM) and G (100 ng/mL) were added during the culture. The cultures were supplemented with 50 U/mL interleukin-2 (Chiron Benelux B. V., The Netherlands) during the culture. The percentage of CMV pp65-specific CD8+T cells was assessed using a PE-CMV pp65 pentamer (Proimmune) and FITC-anti-CD8 mAb. The percentage of Vγ9γδT cells among lymphocytes was assessed by anti-Vγ9TCR and anti-CD3 mAbs. The percentage of Va24NKT cells among lymphocytes expanded was assessed by anti-Va24 and Vβ11 mAbs. The percentage of FoxP3+cells among CD4+CD25+lymphocytes expanded was assessed by anti-FoxP3 mAbs previously described [10].
Patient 1 received seven intradermal (id) injections of 5 X 106 IFN-DCs pulsed with tumor lysates and Z plus G per vaccine at 1 week intervals.
RESULTS AND DISCUSSION
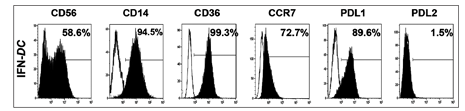
The cells defined as IFN-DCs in this study were CD56++, CD14+++, CD36+++, CCR7++, programmed death ligand (PDL)1++, and PDL2- as shown in Figure 1. The IFN-DCs were also HLA-class I+++, HLA-DR+++, CD80+++, CD86+++, CD40+++, CD54+++, and CD11c+++ (data not shown). CD36 is an important molecule for cell processing [11] and CCR7 is an important molecule for migration of DCs to lymph nodes where interact with lymphocytes [12]. Following treatment with tumor lysates, expression of CCR7 on IFN-DCs increased (data not shown). Note that high expression of CD36 on IFN-DCs treated with tumor lysates is favorable for processing tumor lysates, resulted in the higher induction of CMV pp65-specific CD8+T cells which may be associated with a better clinical outcome.
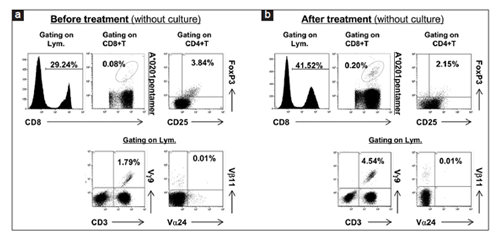
Following DC therapy using IFN-DCs pulsed with tumor lysates and Z plus G for patient 1, increase of CMV pp65-specific CD8+T cells among PB CD8+T cells (from 0.08% to 0.20%) and decrease of CD4+CD25+FoxP3+Treg cells among PB lymphocytes (from 3.84% to 2.15%) were observed on day 30 following seven id injections of IFN-DCs pulsed with the autologous tumor lysates and Z plus G, as shown in Figures 2a and b. In addition, increase of Vγ9γδT cells among lymphocytes expanded was also observed (from 1.79% to 4.54%). However, the percentage of Va24NKT cells among PB lymphocytes was not changed before and after DC therapy at least on day 30 after seven treatments using tumor lysate-pulsed IFN-DCs presumably due to the very low frequency of Va24NKT cells among PB lymphocytes (0.01%) before treatment [Figures 2a]. Note that slightly decrease of CD4+T cells (from 38.40% to 29.65%) and slightly increase of CD3-CD56+NK cells among PB lymphocytes (from 19.88% to 22.8%) were observed on day 30 following seven treatments using tumor lysate-pulsed IFN-DCs copulsed with Z plus G. Taken together, these results show that administration of IFN-DCs pulsed with autologous tumor lysates copulsed with Z plus G for patient 1 induced the enhancements of CMV pp65-specific CD8+T cells and Vγ9γδT cells and the reduction of Treg cells in vivo.
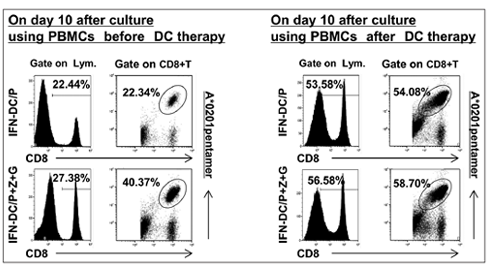
Furthermore, we have compared the expansion ability of CMV pp65- specific CD8+T cells by IFN-DCs pulsed with CMV pp65 peptide versus that by IFN-DCs pulsed with CMV pp65 peptide and Z plus G before DC therapy and after DC therapy in vitro [Figure 3]. In the case of patient 1, HLA-A*0201pentermer positive cells among CD8+T cells expanded by IFN-DCs pulsed with CMV pp65 peptide using PBMCs before DC therapy versus that expanded by IFN- DCs pulsed with CMV pp65 peptide using PBMCs after DC therapy were 22.34% versus 54.08%. Further, HLA-A*0201 pentermer positive cells among CD8+T cells expanded by IFN-DCs pulsed with CMV pp65 peptide and Z plus G before DC therapy versus that expanded by IFN-DCs pulsed with CMV pp65 peptide and Z plus G after DC therapy were 40.37% versus 58.70%. These results indicate that the expansion ability of CMV pp65- specific CD8+T cells by IFN-DCs pulsed with CMV pp65 peptide and Z plus G was higher than that by IFN-DCs pulsed with CMV pp65 peptide alone in both cases, before DC therapy and after DC therapy among which the expansion ability of CMV pp65-specific CD8+T cells by IFN-DCs pulsed with CMV pp65 peptide and Z plus G using PBMCs after DC therapy was highest. Note that the total number of lymphocytes expanded by IFN-DCs/P+Z+G was larger than that expanded by IFN -DCs/P following 10-day culture (i.e., 2.0 X 106 on day 0 and 7.2 X 106 following culture with P alone versus 9.9 X 106 following culture with P+Z+G on day 10).
In humans, most of our knowledge on the specificity and biological role of γδT cells are derived from the analysis of a major peripheral subset referred to as Vγ9Vδ2T cells, comprising 5-10% of all circulating T cell in healthy donors (HDs) [13]. However, the percentage of Vγ9Vδ2T cells among PB lymphocytes from patients with cancer is very low, < 5% [14,15]. In our own observation, the percentage of Vγ9Vδ2T cells among PB lymphocytes from patients with GBM was 1.78 ± 0.16% (n = 4). In this study, following administration of IFN-DCs pulsed with tumor lysates and Z plus G, the percentage of Vγ9γδT cells increased from 1.79% to 4.54%, indicating that following infusions of IFN- DCs pulsed with Z and G, the percentage of Vγ9γδT cells among PB lymphocytes from patient 1 closed to the percentage of Vγ9γδT cells among PB lymphocytes from HDs. Further, it has been shown that Vγ9γδT cells present tumor antigens to aβT cells [16,17]. Thus, it is conceivable that Vγ9γδT cells activated in vivo by infused IFN-DCs pulsed with tumor lysates and Z plus G might play a role as antigen presenting cells in the uptake of apoptotic bodies from tumor cells in vivo, resulted in the increase of CMV pp65-specific CD8+T cell induction. Taken together, administration of tumor lysate-pulsed IFN-DCs copulsed with Z and G is a promising immunotherapy for patients with GBM although more intensive investigations are warranted.
M. Y. analyzed the data, wrote the manuscript, and prepared figures; H. T. designed the experiments; Y. E. performed the experiments, analyzed the data, and prepared figures; X. D. designed the experiments; and M. N. conceived the study, performed the experiments, analyzed the data, and wrote the manuscript.
REFERENCES
- Ohgaki H, Dessen P, Jourde B, Horstmann S, Nishikawa T, Di Patre PL, et al. Genetic pathways to glioblastoma: A population-based study. Cancer Res 2004;64:6892-9.
- Stupp R, Dietrich PY, Ostermann Kraljevic S, Pica A, Maillard I, Maeder P, et al. Promising survival for patients with newly diagnosed glioblastoma multiforme treated with concomitant radiation plus temozolomide followed by adjuvant temozolomide. J Clin Oncol 2002;20:1375-82.
- Lacroix M, Abi-Said D, Fourney DR, Gokaslan ZL, Shi W, DeMonte F, et al. A multivariate analysis of 416 patients with glioblastoma multiforme: Prognosis, extent of resection, and survival. J Neurosurg 2001;95:190-8.
- Prins RM, Cloughesy TF, Liau LM. Cytomegalovirus immunity after vaccination with autologous glioblastoma lysate. N Engl J Med 2008;359:539-41.
- Sabado RL, Bhardwaj N. Dendritic cell vaccines on the move. Nature 2015;519:300-1.
- Cobbs CS, Harkins L, Samanta M, Gillespie GY, Bharara S, King PH, et al. Human cytomegalovirus infection and expression in human malignant glioma. Cancer Res 2002;62:3347-50.
- Mitchell DA, Xie W, Schmittling R, Learn C, Friedman A, McLendon RE, et al. Sensitive detection of human cytomegalovirus in tumors and peripheral blood of patients diagnosed with glioblastoma. Neuro Oncol 2008;10:10-8.
- Eiraku Y, Terunuma H, Yagi M, Deng X, Nicol AJ, Nieda M, et al. Dendritic cells cross-talk with tumour antigen-specific CD8+T cells, Vγ9γδT cells and Va24NKT cells in patients with glioblastoma multiforme and in healthy donors. Clin Exp Immunol 2018;194:54-66.
- Nestle FO, Alijagic S, Gilliet M, Sun Y, Grabbe S, Dummer R, et al. Vaccination of melanoma patients with peptide-or tumor lysate-pulsed dendritic cells. Nat Med 1998;4:328-32.
- Speiser DE, Ho PC, Verdeil G. Regulatory circuits of T cell function in cancer. Nat Rev Immunol 2016;10:599-611.
- Stuart LM, Deng J, Silver JM, Takahashi K, Tseng AA, Hennessy EJ, et al. Response to Staphylococcus aureus requires CD36-mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. J Cell Biol 2005;170:477-85.
- Worbs T, Hammerschmidt SI, Forster R. Dendritic cell migration in health and disease. Nat Rev Immunol 2017;17:30-48.
- Kabelitz D, Wesch D, Pitters E, Zoller M. Potential of human gammadelta T lymphocytes for immunotherapy of cancer. Int J Cancer 2004;112:727-32.
- Abe Y, Muto M, Nieda M, Nakagawa Y, Nicol A, Kaneko T, et al. Clinical and immunological evaluation of zoledronate-activated vgamma9gammadelta T-cell-based immunotherapy for patients with multiple myeloma. Exp Hematol 2009;37:956-68.
- Nakajima J, Murakawa T, Fukami T, Goto S, Kaneko T, Yoshida Y, et al. A phase I study of adoptive immunotherapy for recurrent non-small-cell lung cancer patients with autologous gammadelta T cells. Eur J Cardiothorac Surg 2010;37:1191-7.
- Brandes M, Willimann K, Moser B. Professional antigen-presentation function by human gammadelta T cells. Science 2005;309:264-8.
- Muto M, Baghdadi M, Maekawa R, Wada H, Seino K. Myeloid molecular characteristics of human γd T cells support their acquisition of tumor antigen-presenting capacity. Cancer Immunol Immunother 2015;64:941-9.