Availability of Ag+ Ions Having Most Highly Bactericidal Activity to Suppression by Inhibitions of Viral and Cancerous Cell Growth
Tsuneo Ishida
Citation :Ishida T. Availability of Ag+ ions having most highly bactericidal activity to suppression by inhibitions of viral and cancerous cell growth. Clin Res Immunol 2018;1(2):1-11.
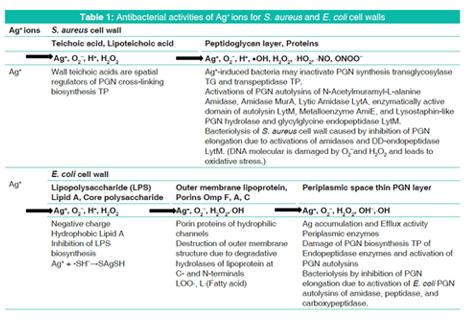
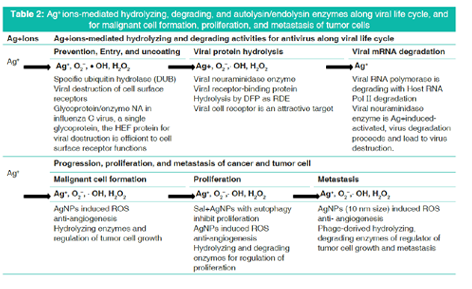
Bacteriolysis against Staphylococcus aureus peptidoglycan (PGN) cell wall by Ag+ ions is due to inhibition of PGN elongation caused by regulation of PGN synthetic transglycosylase and transpeptidase (TP), and enhancement of the activation of PGN autolysins of amidases. The other bacteriolysis and destruction against Escherichia coli cell wall by Ag+ ions are caused by the destruction of outer membrane structure due to degradative enzymes of lipoproteins (LP) at N- and C-terminals and by the inhibition of PGN elongation due to inactivation of PGN TP synthetic enzyme endopeptidase and enhancement of the activations of PGN hydrolases and autolysins of amidase, peptidase, and carboxypeptidase. Ag+ ions-mediated hydrolyzing and degrading activity for antivirus functions by glycosyl hydrolase, viral neuraminidase (NA)/hemagglutinin, cell surface receptor, and the viral degradation lead to virus destructions. Ag+ ions-mediated hydrolyzing activity for anticancer occurs by AgNPs of small size, autophage, and acid hydrolase amidases, and these results lead to the cause of tumor cell growth regulation or cancerous cell death. Silver ions induced ROS generations to occur such as O2-, H2O2,• OH, and OH- producing in the bacterial cell wall, host-virus, and cancerous cell, and lead to oxidative stress, and DNA damages. DNA damages may be due to linear coordinated Ag+ ion complex formations by Ag+ substitution within double and triple hydrogen bonds in DNA base pairs.
Bacteriolysis, DNA base-pairs, hydrolase and degradation, PGN synthesis and autolysin, reactive oxygen species, silver(I) ions
INTRODUCTION
The high antibacterial effects of silver(I) ions such as silver salts and silver nanoparticles (AgNPs) have been widely noticed, should be sufficient to control the majority of the bacterial pathogen, and are utilized as chemotherapy agents. Particularly, AgNPs are associated with immune systems and either act as immunostimulants or immunosuppressors [1]. Released biologically active Ag+ shows a strong affinity for sulphydryl groups and other anionic ligands of proteins, cell membranes, and debris that Ag+ binds protein residues on cell membranes of sensitive bacteria and is absorbed intracellularly by pinocytosis of concentration of 60 ppm Ag+ which Ag+ binds strongly to metallothionein (MT), albumins, and macroglobulins and is metabolized to all tissues other than the brain and the central nervous system [2]. The most notable adverse health effects are argyria and/or argyrosis that additional studies would most likely come to the same conclusions, that is, metallic silver has minimal effect on the human body and soluble silver compounds are likely to produce argyria and argyrosis; therefore, separate recommended permissible exposure limit should be established [3].
Bactericidal activity for Ag+ ions is apparent to be mostly higher from the experimental data that minimum inhibitory concentration (MIC) = 8 ppm and 5 mg/L, minimum bactericidal concentration (MBC) = 32 ppm and 20 mg/L, respectively, for silver sulfate solutions [4] and for AgNPs [5] against S. aureus are obtained that the other, MIC = 4.7 ppm and 40 mg/L, and MBC = 37.5 ppm and 60 mg/L, respectively, for silver nitrate solutions [6] and for AgNPs[7] against E. coli are gained. Silver exists as silver metal and silver ions of different oxidation states of +1, +2, +3, and +4 that the most common states of silver are silver(0) metal and silver(I) ion and both of them interact with thiols in no redox reaction involved that Ag2+, Ag3+, and Ag4+ form state are not of relevance for aqueous solutions and under environmental and biological conditions [8]. Recently, with proceeding development in nanotechnology, AgNPs call attention to potential treatments such as food storage by broad antibacterial effects, prevention of serious diseases, and medical applications [9]. The toxicity of AgNPs is mainly due to release to free silver ions. On the other hand, the antibacterial activity and mechanism of action have been gradually clarified that silver ions may cause Staphylococcus aureus and Escherichia coli bacteria to reach an active but nonculturable state and eventually die, and also have been indicated to the mechanism of inactivation of pathogens by damages and destruction of the bacterial cell membrane [10,11]. The high antibacterial activity factor of Cu2+ and Zn2+ ions may be thought to be caused by binding bacterial surface proteins, cell membrane, and metal-binding complex formations [12]. However, bactericidal elucidation by metal-binding enzyme degradation due to inhibition of peptidoglycan (PGN) elongation and relationships between PGN synthesis and PGN hydrolase/autolysin has been still unclear.
In this review, first, antibacterial effect of silver(I) ions could been elucidated with the attention of PGN growth due to inhibition of PGN syntheses and activation of PGN hydrolases/ autolysins against Gram-positive S. aureus thick PGN layer cell wall and Gram-negative E. coli outer membrane-connecting thin PGN layer cell wall. Second, Ag+ ions-mediated hydrolyzing and degrading functions are treated on glycosyl hydrolase, viral neuraminidase (NA)/hemagglutinin (HA), cell surface receptor for virus degradation of leading to virus destructions, and on AgNPs, autophage, and acid hydrolase amidase for the cancerous cell of leading to cause of the cell death. Finally, Ag+ ions-induced reactive oxygen species (ROS) production is discussed about the lead to oxidative stress and DNA damages against S. aureus, E. coli, and host-viral and cancerous cells, incident to additional DNA damages by Ag+-DNA base pairs interactions.
THE SURFACE ENVELOPE CELL STRUCTURE FOR S. AUREUS AND E. COLI
S. aureus surface layer consists of teichoic acids, lipoteichoic acids, and thick PGN envelope cell wall [13]. There are action sites of transglycosylase (TG) and transpeptidase (TP) mainly on thick PGN layer. The TG is the synthetic enzymes of N-acetylglucosamidase cleavage between N-acetyl glucosamine (NAG) and N-acetylmuramic acid (NAM), and N-acetylmuramidase cleavage between NAM and NAG on glycan chain. The TP is the synthetic enzyme cleavage between Glycine and D-alanine on PGN crosslinking. The other, there are PGN autolysins of N- acetyl-muramyl-L-alanine amidase cleavage, DD-endopeptidases cleavages between glycine and glycine on pentaglycine (Gly)5, and in addition, lysostaphin cleavage between glycine and glycine on PGN cross-linking [14]. PGN synthesis requires glycosyltransferases to polymerize the glycan chain and DD-TP to crosslink the peptides, and PGN cleavage of PGN hydrolases and autolysins is required for reductive cell division and cell separation that amidase has a prominent role in septum cleavage, but lytic TGs and endopeptidases contribute to cell separation, and their role is probably understated due to their greater redundancy, in which they play a key role in regulating cell wall growth as well as other lysis phenomena [15].
On the other hand, E. coli cell wall consists of lipid A, lipopolysaccharide (LPS), porin proteins, outer membrane of lipoprotein, and thinner 2-7 nm PGN layer in 30-70 nm periplasmic space [13]. Degradative enzymes of lipoproteins at N- and C-terminals are both Endopeptidase between phospholipid-Lipoprotein bond and Amidase between L-Ala-NAM bond via E. coli outer membrane, within from lipoprotein to PGN. It is apparent that these degradative enzyme hydrolases lead to destruction of E. coli outer membrane structure.
In the molecular bonding manner of E. coli cell wall and peri-plasmic PGN, there are E. coli PGN synthetic enzymes TG of Glucosaminidase cleavage, Muramidase cleavage on glycan chain, and TP of Endopeptidase cleavage on cross-linking. The other, PGN hydrolases and autolysins are degradative enzymes of Amidase cleavage, Peptidase cleavage, and Carboxypeptidase cleavage [6]. Penicillin-binding protein 2 and RodA (encoded downstream of the PBP2 gene) are required for the PGN synthesis of glycan strands during elongation and the periplasmic amidase cleave only one-sixth of the PGN that is turned over by the lytic TGs. E. coli has at least 13 periplasmic PGN hydrolases (autolysins), which can collectively cleave almost any glycoside and amide bond [14]. Interactions of PGN molecular structure with PGN syntheses and PGN autolysins influence, in any event, the bacteriolytic cell walls.
AG+ IONS-MEDIATED VIRUS AND CANCER/TUMOR HYDROLYZING AND DEGRADING MECHANISM
Silver(I) ions reactive with thiol, and then generates silver(I) thiolate compounds. Silver ion complexes with both inorganic and organic thiols with redox reaction involved that with inorganic thiols such as HS- and S2+, it is possible to form many species such as AgSH, [Ag(SH)2]- , and [Ag2(SH)2S]2-depending on the concentration of the anions present [15].
Ag+ + (-SH)- → AgSH → [Ag2(SH)2S]2-
Tol protein (Tol)- protein-associated lipoprotein (Pal) system is composed of five proteins that TolA, TolQ, and TolR are inner membrane proteins, TolB is a periplasmic protein, and Pal, the peptidoglycan-associated LP is anchored to the outer membrane [16]. Ag+ ions induced Tol-Pal complex is antimicrobial agents widely used, it has recently been demonstrated to be essential for bacterial survival and pathogenesis that outer membrane structure may be destroyed. It is unclear whether both Amidase and Endopeptidase of LP at C- and N-terminals are simultaneously activated by Ag+ ions, and whether virus and tumor cell-hydrolyzing and degrading functions are occurred by predominant activation of Ag+-amidase reaction.
DISCUSSION
There is necessarily required for the adequate balance between PGN synthesis and PGN autolysin to growth of S. aureus PGN cell wall that when the balance is lost to be imbalanced. Bacteriolysis and destruction of the cell wall should be occurred. The bacteriolysis of S. aureus PGN cell wall by silver ions is caused by inhibition of PGN elongation due to inactivation of PGN synthesis and enhancement of activation of PGN autolysins, with the similar bacteriolytic phenomenon for E. coli cell wall [14].
The released Ag+ ions that penetrated from AgNPs into bacterial cells can inhibit the growth of Gram-positive B. subtilis bacterium which exerts toxicity by damaging cellular membrane, degrading chromosomal DNA, lowering reductase activity, and reducing protein expression [17]. Wall teichoic acids are spatial regulators of PGN cross-linking biosynthesis TP,[18] and silver ions could inhibit both TG and TP enzymes of the PGN that Ag+-induced bacteria may inactivate PGN synthesis TG [19] and TP [20,21]. Lysostaphin-like PGN hydrolase and glycylglycine endopeptidase LytM may function as TP enzyme [22].
Ag+ iduced amidase of S. aureus PGN autolysin Lytic activity was inhibited by glucosamine, NAG, Hg2+, Fe3+, and Ag+,[23] and Ag+ binding Rv3717 showed no activity on polymerized PGN and but, it is induced to a potential role of N-Acetylmuramyl-L-alanine Amidase,[24] PGN murein hydrolase activity and generalized autolysis; Amidase MurA,[25] Lytic Amidase LytA,[26] enzymatically active domain of autolysin LytM,[27] metal-dependent metalloenzyme AmiE [28] as prevention of the pathogen growth. The activations of these PGN autolysins could be enhanced the inhibitions of PGN elongation simultaneously, with bacteriolysis of S. aureus PGN cell wall. Bacteriolysis of S. aureus PGN cell wall by Ag+ ions is caused by inhibition of PGN elongation due to inactivation of PGN TG or TP and enhancement of activation of PGN autolysins of amidases. O2- and H2O2 permeate into membrane and cytoplasm, and then, DNA molecular is damaged by oxidative stress [29].
E. coli cell wall is comprised LPS, LP, and PGN as a thinner layer within periplasmic space. When permeability of silver ions in the E. coli cell wall, highly anionic LPS with hydrophobic lipid A, core polysaccharide, O-polysaccharide, is liable to be explosive, inhibition of LPS biosynthesis may be possibility to occur by active hydrolases [30]. The OmpA, OmpC, and OmpF porins of LP have metallic cation selective and hydrophilic membrane crossing pore, to be effective for silver transfer [31]. Age-resistant mutants of E. coli display active efflux of Ag+ and are deficient in porins that active efflux may play a major role in silver resistance, which is likely to be enhanced synergistically by decreases in OM permeability [32]. Physicochemical interaction of E. coli cell envelopes suggested that the adsorption of the cell wall or envelope to clay has masked or neutralized chemically reactive adsorption sites normally available to metal ions that metal binding capacity of metal cation bridging in isolated envelopes was determined by atomic absorption spectroscopy[33]. Silver adsorption by E. coli cells displays MTs anchored to the outer membrane protein LamB that the complete MT sequences are anchored by their N-termini and C-termini to the permissive site 153 of the protein [34]. Recently, Ag+ ions into E. coli cell wall are elucidated to be occurred E. coli under ionic silver stress which Ag+-dependent regulation of gene expression is TP acting on the structural integrity of the cell wall [35].
Tol protein (Tol)- Pal system is composed of five proteins that TolA, TolQ, and TolR are inner membrane proteins, TolB is a periplasmic protein, and Pal, the peptidoglycan-associated LP, is anchored to the outer membrane [16]. Ag+ ions induced Tol-Pal complex is antimicrobial agents widely used; it has recently been demonstrated to be essential for bacterial survival and pathogenesis that outer membrane structure may be destroyed [36,37]. It is unclear whether both Amidase and Endopeptidase of LP at C- and N-terminals are simultaneously activated by Ag+ ions. However, outer membrane may be probably destroyed by predominant activation of LP -amidase.
Silver ions may be accumulated in E. coli periplasmic space, in which the silver ions are spent to the activation of bacteriolysis of the cell wall and efflux activity to extracellular cell. Then, LP -endopeptidase may be degradative by Ag+ binding proteins [38]. The other, it is unclear that the silver-induced PGN biosyntheses TG/TP should be inhibited by the silver ions [39-41]. However, silver ions inactivate TP of endopeptidase by due to the destructive observation of bacterial cell walls [35]. Silver ions could activate E. coli PGN autolysins of amidase, peptidase, and carboxypeptidase,[42,43]. such as silver depending PGN autolysin, AmiC,[44] AmiD,[45] Muramidase,[23] amino acid amidase,[46] Carboxypeptidase A,[47] Amidase zinc-containing amidase; AmpD,[48] zinc-present PGLYRPs,[49] Carboxypeptidase-degraded aldolase,[50] and CarboxypeptidaseY [51] serve to be effective for the PGN autolysins. It is particularly worth noting that enhancement of the activities of autolysins is characterized on PGN carboxypeptidase and TP-endopeptidase [42] requiring divalent cations. Accordingly, the inhibition of PGN elongation had occurred by silver ion induced activities of PGN hydrolases and autolysins.
From above-mentioned results, antibacterial activities of Ag+ ions for bacteriolytic process of S. aureus PGN and E. coli cell walls are shown in Table 1.
Bacterial lysozymes were first glycosyl hydrolases to have their three-dimensional structures solved that the two catalytic amino acids were identified as aspartate and glutamate residues which in most glycosyl hydrolases studied since only aspartate and/or glutamate residues have been found to perform catalysis [52]. Glycoside hydrolases are enzymes that catalyze the hydrolysis of the glycosidic linkage of glycosides, leading to the formation of a sugar hemiacetal or hemiketal and the corresponding free aglycon.
Recent example is viral NA and bacterial sialidase, where the transition state is thought to be stabilized with the help of a tyrosine. NA enzymes are glycoside hydrolase enzymes that cleave the glycosidic linkages of neuraminic acids. The NA is important for the initiation, release of influenza virus infection in human airway epithelium that influenza virus NA plays an essential role in release and spread of progeny virions, intracellular viral replication cycle which viral NA provide an important role early in infection and further rationale for the prophylactic use of NA inhibitors [53]. The structure of the influence virus NA, the special organization of their active site, the mechanism of carbohydrate chains desialylation by NA, and its role in the influenza virus function at different stages of the viral infectious cycle are considered that the NA substrate specificity and different approaches relating the activity of this enzyme are investigated and are provided that along with considerations on the mechanisms of resistance of modern influenza viruses to those antivirals [54]. Further, NA protein from influenza A virus functions to promote the viral release and is one of major surface antigens that the receptor-destroying activity in NA resides in the distal head domain that is linked to the viral membrane by an N-terminal hydrophobic transmembrane domain (TMD) [16] The NA TMD in H1N1 viruses has become more polar to maintain compatibility with the evolving enzymatic head domain [55]. The interaction of influenza A virus glycoproteins with cell surface receptors is a major determinant of infectivity and therefore transmissibility that factors are necessary to determine pandemic potential which influenza A viruses generally mediate binding to cell surface sialic acid receptors through the HA) glycoprotein, with the NA. NA glycoprotein being responsible for cleaving the receptor to allow virus release [56]. This N9 NA has an active Hb site which binds to sialic acid, which enhances overall virus binding to sialic acid receptor analog. The N9 NA can also contribute to receptor binding due to unusual kinetic characteristics of the sialidase site which specifically enhance binding to human-like α2,6-linked sialic acid receptor. Viral NA can be assumed that the NA enzyme is corresponded to enzyme of bacterial PGN autolysin. When the viral NA enzyme is activated, and virus protein homeostasis is lost, virus degradation proceeds, in which virus lead to apoptotic death and the virus is destroyed. Ag+ ions induced viral NA activations are enhanced in action sites of virus,[57]. subsequently, the virus growth is suppressed, and virus destruction occurs. Turnip yellow mosaic virus (TYMV) encodes a precursor replication polyprotein that is processed by the endoproteolytic activity of its internal cysteine proteinase domain (PRO) which PRO is actually a multifunctional enzyme with a specific ubiquitin hydrolase (DUB) activity that contributes to viral infectivity [58]. Thus, DUBs can proceed to specifically hydrolyze under Ag+ ions-induced state both iso- and endopeptide bonds with different sequences that were shed light on how such a compact protein achieves a diversity of key functions in viral genome replication and host-pathogen interaction. Influenza viruses induced a host shut off mechanism leading to the general inhibition of host gene expression in infected cells. The large subunit of host RNA polymerase II (Pol II) is degraded in infected cells which this degradation is mediated by the viral RNA polymerase that associated with Pol II, in which increased ubiquitylation of Pol II in infected cells and on the expression of the viral RNA polymerase suggesting that the proteasome pathway plays a role in Pol II degradation and further, expression of the viral RNA polymerase results in the inhibition of Pol II transcription [59].
Viral destruction of cell surface receptors, in general, the glycoproteins of several lipid-enveloped viruses, including orth-myxoviruses (influenza A, B, and C), toroviruses, and coronaviruses, have three important functions: To recognize the receptor on the cell surface, to mediate viral fusion with the cell membrane, and to destroy the receptor [60]. In highly infections influenza A and B viruses, the receptor-binding and membrane-fusion activities of cell entry carried out by the glycoprotein HA that the receptor-destroying enzyme (RDE) activity important for virus release is conducted by the glycoprotein/enzyme NA. In influenza C virus, a single glycoprotein, the hemagglutinin-esterase-fusion (HEF) protein, processes all three functions [60]. Thus, the structures of HA, HFE, and HE should serve as a basis for the understanding of how viruses recognize their receptors on the cell surface, mediate viral fusion with the cell membrane, and then destroy their receptors. H5Nx avian influenza viruses have a strict avian receptor binding preference, while recombinantly expressed NAs are sensitive to FDA-approved and investigational antivirals [61]. Sendai virus (SeV) particles could not attach to or penetrate the infected cells, and the hemagglutinin-neuraminidase protein of SeV was involved in the interference, in which the α2,3-linked sialic acids were specifically reduced in the SeV-infected cells, but the level of α2,6-linked sialic acids had not changed [62]. These results provide concrete evidence of destruction of the specific SeV receptor of sialic acid, being relevant to homologous interference by SeV. The hemagglutinin-esterases (HEs) are a family of viral envelop glycoproteins that mediate reversible attachment to O-acetylated sialic acids by acting both as lectins and as RDEs. Coronavirus (CoV) HE arose from an influenza C-like HEF that the HE receptor-binding domain underwent remodeling to such extent that the ligand is now bound in opposite orientation [63]. Thus, the CoV HEs structure offers insight into corona and influenza virus evolution.
Inhibition of influenza C virus infection was observed only at an inhibitor concentration that was about 100-fold higher than the maximum concentration of inhibitor that could be inactivated by the RDE of a given amount of virus [64]. RDE is not required to inactivate inhibitors. The infectious salmon anemia virus (ISAV) acetylesterase activity was inhibited by di-isopropyl fluorophosphate (DFP) in dose-dependent fashion but not by other known hydrolase inhibitors, suggesting that a serine residue is part of the active site, in which the effect of DFP on agglutination/ elution of erythrocytes by ISAV demonstrated that the acetylesterase activity is the bona fide RDE [65]. The viral cell receptor is an attractive target for anti-viral strategies that antibodies significantly inhibited (avian sarcoma/ leukosis virus including the A-J ten subgroups) infection and replication which cell lines with the expression of viral receptor-binding protein could be as efficient tools for isolating functional receptors to identify novel anti-viral targets [66].
AgNPs possess unique cytotoxic features that AgNPs with size of 5 nm and 35 nm can are kill osteosarcoma cells independently from their actual p53 status and induce p53-independent cancer cell apoptosis [67]. AgNPs with 2.6 and 18 nm size decreased viability, proliferation and caused the death of pancreatic cancer cells in size- and concentration-dependent manner that cellular uptake of AgNPs resulted in apoptosis, autophagy, necroptosis, and mitotic catastrophe [68]. AgNPs also have an anticancer activity of endoplasmic reticulum stress, oxidative stress, and mitochondrial impairment triggering cell death by apoptosis and autophagy activation [69]. Engineered nanoparticles (ENPs) with sizes no >100 nm are able to enter the human body and accumulate in organs such as brain, liver, lung, testes, and cause toxic effects, and ENPs cytotoxicity in living cells, sizes, shapes, surface charges, agglomeration status all play a deciding ENP safety and suitability for such roles, in which ENPs induce cell apoptosis with potential cancer therapy [70]. Hydrolase activity is an essential regulator of growth of cancer and tumor cell that the enzyme activity drives the functional contribution of this protein to tumor-cell growth which retinoblastoma-binding protein 9 (RBBP9) as a tumor-associated serine hydrolase that displays elevated activity in pancreatic carcinomas [71]. Thus, RBBP9-mediated suppression of TGF-β signaling is required for E-cadherin expression as loss of the serine hydrolase activity leads to a reduction in E-cadherin levels and a concomitant decrease in the integrity of tumor cell-cell junctions [71]. The hydrolases have enzyme-mediated hydrolytic activation that plays the degradative role of the tumor cell. As the main compartment for intracellular degradation and subsequent recycling of cellular constituents, the lysosomes receive both hetero- and auto-phagic cargo, which in the degradative lumen of this organelle find their final destination. The degradation is carried out by many acid hydrolases such as phosphatase, nucleases, glycosidases, peptidases, sulfatases, and lipases capable of digesting all major cellular macromolecules [72]. The physical disruption including leakage of lysosomal hydrolases into the cytosol, led to programmed cell death, and necrosis. Numerous regulatory proteins direct the addition of ubiquitin to lysine residues on target proteins, and there are countered by an army of deubiquitinating enzymes. BRCA1-associated Protein-1 helps to control cell proliferation by resulting host cell factor-1 protein levels and by associating with a gene involved in the G1-S transition [73]. Therefore, silver ions induced carboxy-terminal hydrolase activation may be promoted the apoptosis and the necrosis of cancer cells. However, Ag+ ion-mediated hydrolyzing and degrading method is incompletely yet established without information for Ag+ ions-associated enzymes. Ubiquitin C-terminal hydrolase-L1 (UCH-L1) increases cellular ROS levels and promote tumor invasion, in which silencing UCH-L1, as well as inhibition of H2O2 generation by catalase, a NOX inhibitor, suppressed the migration potential of B16F cells, indicating that UCH-L1 promotes cell migration by up-regulating H2O2 generation [74]. The other, gold nanorod induced apoptosis specifically in cancer cells by affecting lysosome and mitochondria [75]. Lysosomes are membrane-bound organelles containing that function in the degradation of macromolecules delivered through the endocytic, phagocytic, and autophagic pathways, in which AgNPs induced cell death was increased by bafilomycin Al treatment, and the perturbation of lysosomal pH by AgNP exposure may play a role in AgNP agglomeration and subsequent cellular damage in cancer cell and moderate lysosomal permeabilization can result in apoptosis or apoptosis-like cell death [76].
Elongated nanoparticle aggregates and generated hundreds of pico Newton to dramatically damage the plasma and lysosomal membranes, whereas the physical disruption, including leakage of lysosomal hydrolases into the cytosol, led to programmed cell death and necrosis [77]. Accordingly, the perturbation of lysosomal pH by AgNP exposure may play a role in AgNP agglomeration and subsequent cellular damage in A549 cells. The microbial amidases enzymes are in great demand for use as therapeutic agents against many dreadful diseases that in the presence of these enzymes, the tumor cells failure to survive which L-aspaginase and L-glutaminase can use as potent antitumor or antileukemic drugs [78]. Identification of N-acylethanolamine-hydrolyzing acid amidase inhibitors has been able to reduce cell proliferation and migration and cause cell death on different bladder cancer cell lines [79]. Further, the use of antibody-enzyme conjugates for cancer therapy [80] that LytA-like N-acetylmuramoyl-L-lanine amidases [81] such as lytic amidase LytA,[26] LytA autolysin,[82] autolysin LytA,[83] and autophagy,[84] and murein lytic cleavage agents on glycan-chain-like as chimeric phage endolysin such as Ply187[85] and LysPA26 [86] in cancer therapy. These works provide a novel strategy of designing magnetic nanomedicines for mechanical destruction of cancer cells. Comparing with bacterial PGN autolysins, the research of cancer cell hydrolases will become important as a significant advancement for cancer and tumor cells in the future.
Table 2 shows Ag+ ions-mediated hydrolyzing, degrading, and autolysin/endolysin enzymes for antiviral life cycle, and anticancer of malignant cell formation, proliferation, and metastasis of tumor cells.
PRODUCTION OF ROS, OXIDATIVE STRESS DAMAGE, AND KILLING AGAINST S. AUREUS, E. COLI, AND VIRAL HOST/CANCEROUS CELLS
Silver ions reacted with -SH and H+ generates. In E. coli, free radicals O2-, OH-, •OH, and H2O2 are formed as follows.[67]
For the penetration of Ag+ ions to PGN cell wall of S. aureus, the ROS production such as superoxide anion radical O2-, hydroxyl radical •OH, hydrogen peroxide H2O2 occurred from superoxide radical O2- molecular[87]. For viral and tumorous cells also, Ag+ induced ROS and H2O2 generation lead to oxidative stress and DNA damage [68].
2O2- + 2H+ → H 2 O2 + O2
O2- + H 2 O2 → OH- + •OH + O2
In the cell wall, reacting with polyunsaturated fatty acids:
LH + OH• → L• + HOH
L• + O2 → LOO•
LH + LOO• → L• + LOOH
Ag+-containing PGN recognition proteins induce ROS production of H2O2, O-, and HO•, and then the ROS and H2O2 occur with the oxidative stress and killing by stress damage [88].
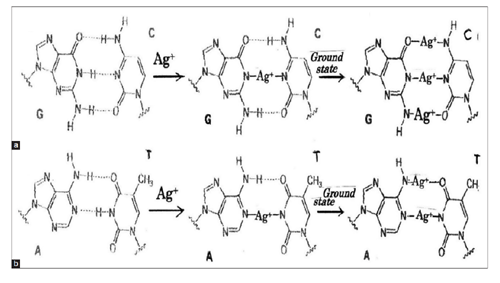
Ag+ ion-induced occurrence of generations of ROS and hydrogen peroxide H2O2 in bacterial cells and DNA, in which formation of DNA damage resulting from a release of catalytic binding of zinc ion to DNA with generation of •OH radicals, and by reaction of H2O2 with the metal produces the strand breaks in DNA as well as DNA base-pairs modifications and deoxyribose fragmentation. Transfer of Ag+ ions into triple and double hydrogen bonds within DNA base-pairing G(guanine)=C(cytosine) and adenine=T (thymine) pairs occurs by Ag+ ion substitution shown in Figure 1. Thus, it may be considered that DNA damages due to Ag+ two-coordinated complex formations within DNA base-pairs of triple hydrogen bond G=C and double hydrogen bond A=T are subjected in the ground state. A=T base pairs are less stable than G=C base pairs in Ag+-DNA reaction [89].
CONCLUSION
- Ag+-induced S. aureus may inactivate PGN synthesis TG and TP. Bacteriolysis of S. aureus PGN cell wall, in which wall teichoic acids control PGN synthesis cross-linking TP, is due to the inhibition of PGN elongation by enhancing the activities of PGN autolysins; amidase AmiA and AmiE, and PGN hydrolase Lysostaphin-like endopeptidase (Glycine-Glycine bond cleavage).
- Bacteriolysis and destruction of E. coli cell wall are due to the damage of LPS synthesis, destructing of outer membrane structure by degrading of LP at C-,N-terminals, due to inhibition of PGN formations by inactivation of carboxypeptidase and TP-endopeptidase, and activities of PGN autolysins of amidase, peptidase, and carboxypeptidase.
- Ag+ ions-mediated hydrolyzing and degrading activities for anti-virus are functioned by glycosyl hydrolase, viral NA/HA, cell surface receptor, and subsequently, virus degradation is led to virus destructions.
- The viral NA enzyme is corresponded to enzyme of bacterial PGN autolysin, and the viral cell receptor is an attractive target for anti-viral strategies as RDE.
- Ag+ ions-associated hydrolysis has enzyme-mediated hydrolytic activation that plays the degradative role of the tumor cell. The intracellular degradation may be carried out by acid hydrolases such as phosphatase, nucleases, glycosidases, peptidases, and sulfatases capable of digesting all major cellular macromolecules. Ag+ ions-induced hydrolyzing and degrading functions for anticancer lead to the cause of tumor cell growth regulation or cancerous cell death.
- By the penetration of silver ion into S. aureus cell wall, production of O2, H+, and H2O2, ONOO- occurs against S. aureus. The other, the productions of O2-, H+ in the outer membrane, and H2O2, OH-, •OH in periplasmic space occur against E. coli cell wall. For Ag+ induced viral and cancerous cell, these ROS and H2O2 give the damages cell membrane proteins and DNA molecular in the cytoplasm.
- DNA damages due to linear coordinated Ag+ ion complex formation within DNA base-pairs of triple hydrogen bond G=C, double hydrogen bond A=T may be occurred in the cytoplasm of bacterial cells.
REFERENCES
- Chakraborty B, Pal R, Ali M, Singh LM, Rahman DS, Kumar Ghosh S, et al. Immunomodulatory properties of silver nanoparticles contribute to anticancer strategy for murine fibrosarcoma. Cell Mol Immunol 2016;13:191-205.
- Lansdown AB. A pharmacological and toxicological profile of silver as an antimicrobial agent in medical devices. Adv Pharmacol Sci 2010;2010:910686.
- Drake PL, Hazelwood KJ. Exposure-related health effects of silver and silver compounds: A review. Ann Occup Hyg 2005;49:575-85.
- Sandstrom S. The Antibacterial Effect of Silver with Different Release Kinetics. Master Science Thesis in Biotechnology. Sweden: Chalmers University and Biological Engineering; 2011.
- Khan A, Shaheen A, Mahmood T, Rehman W, Yaqoob N. Novel synthesis and characterization of silver nanoparticles from leaf aqueous extract of Aloe vera and their anti-microbial activity. J Nanosci Nanotechnol Appl 2017;1:1-9.
- Ishida T. Bacteriolyses of Cu2+ solution on bacterial cell walls/ cell membrane and DNA base pairing damages. Jpn Biomed Res Trace Elements 2016;27:151-61.
- Ruparelia JP, Chatterjee AK, Duttagupta SP, Mukherji S. Strain specificity in antimicrobial activity of silver and copper nanoparticles. Acta Biomater 2008;4:707-16.
- Behra R, Sigg L, Clift MJ, Herzog F, Minghetti M, Johnston B, et al. Bioavailability of silver nanoparticles and ions: From a chemical and biochemical perspective. J R Soc Interface 2013;10:20130396.
- Lindgren AL. The effects of silver nitrate and silver nanoparticles on Chlamydomonas reinhardtii: A proteomic approach. PLoS One 2014;9:1-34.
- Jung WK, Koo HC, Kim KW, Shin S, Kim SH, Park YH, et al. Antibacterial activity and mechanism of action of the silver ion in Staphylococcus aureus and Escherichia coli. Appl Environ Microbiol 2008;74:2171-8.
- Varkey AJ, Dlamini MD, Mansuetus AB, Tiruneh AT. Germicidal action of some metals/metal ions in combating E. coli bacteria in relation to their electro-chemical properties. J Water Resour Protein 2013;5:1132-43.
- Ishida T. Halo-inhibitory zone tests and antibacterial activities for some metallic salts aqueous solutions. Jpn J Prev Med 2017;11:93-9.
- Silhavy TJ, Kahne D, Walker S. The bacterial cell envelope. Cold Spring Harb Perspect Biol 2010;2:a000414.
- Typas A, Banzhaf M, Gross CA, Vollmer W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat Rev Microbiol 2011;10:123-36.
- Shuang TH, Christopher BM, Kristina T, Richard GC. Chemical interactions between silver nanoparticles and thiols: A comparison of mercaptohexanol against cysteine. Sci China Chem 2014;57:1199-210.
- Cascales E, Bernadac A, Gavioli M, Lazzaroni JC, Lloubes R. Pal lipoprotein of Escherichia coli plays a major role in outer membrane integrity. J Bacteriol 2002;184:754-9.
- Hsueh YH, Lin KS, Ke WJ, Hsieh CT, Chiang CL, Tzou DY, et al. The antimicrobial properties of silver nanoparticles in Bacillus subtilis are mediated by released Ag+ ions. PLoS One 2015;10:e0144306.
- Atilano ML, Pereira PM, Yates J, Reed P, Veiga H, Pinho MG, et al. Teichoic acids are temporal and spatial regulators of peptidoglycan cross-linking in Staphylococcus aureus. Proc Natl Acad Sci U S A 2010;107:18991-6.
- Baizman ER, Branstrom AA, Longley CB, Allanson N, Sofia MJ, Gange D, et al. Antibacterial activity of synthetic analogues based on the disaccharide structure of moenomycin, an inhibitor of bacterial transglycosylase. Microbiology 2000;146 Pt 12:3129-40.
- Oka T, Hashizume K, Fujita H. Inhibition of peptidoglycan transpeptidase by beta-lactam antibiotics: Structure-activity relationships. J Antibiot (Tokyo) 1980;33:1357-62.
- Ortiz-Gila MA, Nunez-Anita RE, Srenas-Arrocena M, Omar MA, Joel R, Emiliano R, et al. Silver nanoparticles for the inhibition of S. aureus. Entreciencias 2015;3:133-42.
- Ramadurai L, Lockwood KJ, Nadakavukaren MJ, Jayaswal RK. Characterization of a chromosomally encoded glycylglycine endopeptidase of Staphylococcus aureus. Microbiology 1999; 145 (Pt 4):801-8.
- Croux C, Canard B, Goma G, Soucaille P. Purification and characterization of an extracellular muramidase of Clostridium acetobutylicum ATCC 824 that acts on non-N-acetylated peptidoglycan. Appl Environ Microbiol 1992;58:1075-81.
- Prigozhin DM, Mavrici D, Huizar JP, Vansell HJ, Alber T. Structural and biochemical analyses of Mycobacterium tuberculosis N-acetylmuramyl-L-alanine amidase rv3717 point to a role in peptidoglycan fragment recycling. J Biol Chem 2013;288:31549-55.
- Carroll SA, Hain T, Technow U, Darji A, Pashalidis P, Joseph SW, et al. Identification and characterization of a peptidoglycan hydrolase, murA, of Listeria monocytogenes, a muramidase needed for cell separation. J Bacteriol 2003; 185:6801-8.
- Mellroth P, Sandalova T, Kikhney A, Vilaplana F, Hesek D, Lee M, et al. Structural and functional insights into peptidoglycan access for the lytic amidase lytA of Streptococcus pneumoniae. MBio 2014;5:e01120-13.
- Jagielska E, Chojnacka O, Sabala I. LytM fusion with SH3b-like domain expands its activity to physiological conditions. Microb Drug Resist 2016;22:461-9.
- Zoll S, Patzold B, Schlag M, Gotz F, Kalbacher H, Stehle T, et al. Structural basis of cell wall cleavage by a staphylococcal autolysin. PLoS Pathog 2010;6:e1000807.
- Morina F, Vidovic M. Induction of peroxidase isoforms in the roots of two Verbascum thapsus L. Populations is involved in adaptive responses to excess Zn2+ and Cu2+. Bot Serbica 2015;39:151-8.
- Langley S, Beveridge TJ. Effect of O-side-chain-lipopolysaccharide chemistry on metal binding. Appl Environ Microbiol 1999;65:489-98.
- Blindauer CA. Advances in the molecular understanding of biological zinc transport. Chem Commun (Camb) 2015; 51:4544-63.
- Li XZ, Nikaido H, Williams KE. Silver-resistant mutants of Escherichia coli display active efflux of Ag+ and are deficient in porins. J Bacteriol 1997;179:6127-32.
- Walker SG, Flemming CA, Ferris FG, Beveridge TJ, Bailey GW. Physicochemical interaction of Escherichia coli cell envelopes and Bacillus subtilis cell walls with two clays and ability of the composite to immobilize heavy metals from solution. Appl Environ Microbiol 1989;55:2976-84.
- Sousa C, Kotrba P, Ruml T, Cebolla A, De Lorenzo V. Metalloadsorption by Escherichia coli cells displaying yeast and mammalian metallothioneins anchored to the outer membrane protein lamB. J Bacteriol 1998;180:2280-4.
- Saulou-Berion C, Gonzalez I, Enjalbert B, Audinot JN, Fourquaux I, Jamme F, et al. Escherichia coli under ionic silver stress: An integrative approach to explore transcriptional, physiological and biochemical responses. PLoS One 2015; 10:e0145748.
- Veronesi G, Deniaud A, Gallon T, Jouneau PH, Villanova J, Delangle P, et al. Visualization, quantification and coordination of Ag+ ions released from silver nanoparticles in hepatocytes. Nanoscale 2016;8:17012-21.
- Slavin YN, Asnis J, Hafeli UO, Bach H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J Nanobiotechnology 2017;15:65.
- Godlewska R, Wisniewska K, Pietras Z, Jagusztyn-Krynicka EK. Peptidoglycan-associated lipoprotein (Pal) of gram-negative bacteria: Function, structure, role in pathogenesis and potential application in immunoprophylaxis. FEMS Microbiol Lett 2009;298:1-1.
- Egan AJ, Biboy J, van't Veer I, Breukink E, Vollmer W. Activities and regulation of peptidoglycan synthases. Philos Trans R Soc Lond B Biol Sci 2015;370:Pii: 20150031.
- Singh SK, SaiSree L, Amrutha RN, Reddy M. Three redundant murein endopeptidases catalyse an essential cleavage step in peptidoglycan synthesis of Escherichia coli K12. Mol Microbiol 2012;86:1036-51.
- Ramachandran V, Chandrakala B, Kumar VP, Usha V, Solapure SM, de Sousa SM, et al. Screen for inhibitors of the coupled transglycosylase-transpeptidase of peptidoglycan biosynthesis in Escherichia coli. Antimicrob Agents Chemother 2006;50:1425-32.
- Gonzalez-Leiza SM, de Pedro MA, Ayala JA. AmpH, a bifunctional DD-endopeptidase and DD-carboxypeptidase of Escherichia coli. J Bacteriol 2011;193:6887-94.
- Rivera I, Molina R, Lee M, Mobashery S, Hermoso JA. Orthologous and paralogous ampD peptidoglycan amidases from gram-negative bacteria. Microb Drug Resist 2016;22:470-6.
- Garcia DL, Dillard JP. AmiC functions as an N-acetylmuramyl-l-alanine amidase necessary for cell separation and can promote autolysis in Neisseria gonorrhoeae. J Bacteriol 2006;188:7211-21.
- Pennartz A, Genereux C, Parquet C, Mengin-Lecreulx D, Joris B. Substrate-induced inactivation of the Escherichia coli amiD N-acetylmuramoyl-L-alanine amidase highlights a new strategy to inhibit this class of enzyme. Antimicrob Agents Chemother 2009;53:2991-7.
- Komeda H, Hariyama N, Asano Y. L: -Stereoselective amino acid amidase with broad substrate specificity from Brevundimonas diminuta: Characterization of a new member of the leucine aminopeptidase family. Appl Microbiol Biotechnol 2006;70:412-21.
- Coombs TL, Omote Y, Vallee BL. The zinc-binding groups of carboxypeptidase A. Biochemistry 1964;3:653-62.
- Carrasco-Lopez C, Rojas-Altuve A, Zhang W, Hesek D, Lee M, Barbe S, et al. Crystal structures of bacterial peptidoglycan amidase ampD and an unprecedented activation mechanism. J Biol Chem 2011;286:31714-22.
- Wang M, Liu LH, Wang S, Li X, Lu X, Gupta D, et al. Human peptidoglycan recognition proteins require zinc to kill both gram-positive and gram-negative bacteria and are synergistic with antibacterial peptides. J Immunol 2007;178:3116-25.
- Drechsler ER, Boyer PD, Kowalsky AG. The catalytic activity of carboxypeptidase-degraded aldolase. J Biol Chem 1959;234:2627-34.
- Hayashi R, Bai Y, Hata T. Further confirmation of carboxypeptidase Y as a metal-free enzyme having a reactive serine residue. J Biochem 1975;77:1313-8.
- Davies G, Henrissat B. Structures and mechanisms of glycosyl hydrolases. Structure 1995;3:853-9.
- Matrosovich MN, Matrosovich TY, Gray T, Roberts NA, Klenk HD. Neuraminidase is important for the initiation of influenza virus infection in human airway epithelium. J Virol 2004;78:12665-7.
- Shtyrya YA, Mochalova LV, Bovin NV. Influenza virus neuraminidase: Structure and function. Acta Naturae 2009;1:26-32.
- da Silva DV, Nordholm J, Dou D, Wang H, Rossman JS, Daniels R, et al. The influenza virus neuraminidase protein transmembrane and head domains have coevolved. J Virol 2015; 89:1094-104.
- Benton DJ, Wharton SA, Martin SR, McCauley JW. Role of neuraminidase in influenza A(H7N9) virus receptor binding. J Virol 2017;91:1-10.
- Holmquist L. Activation of vibrio cholerae neuraminidase by divalent cations. FEBS Lett 1975;50:269-71.
- Lombardi C, Ayach M, Beaurepaire L, Chenon M, Andreani J, Guerois R, et al. A compact viral processing proteinase/ ubiquitin hydrolase from the OTU family. PLoS Pathog 2013;9:e1003560.
- Vreede FT, Chan AY, Sharps J, Fodor E. Mechanisms and functional implications of the degradation of host RNA polymerase II in influenza virus infected cells. Virology 2010;396:125-34.
- Mesecar AD, Ratia K. Viral destruction of cell surface receptors. Proc Natl Acad Sci U S A 2008;105:8807-8.
- Yang H, Carney PJ, Mishin VP, Guo Z, Chang JC, Wentworth DE, et al. Molecular characterizations of surface proteins hemagglutinin and neuraminidase from recent H5Nx avian influenza viruses. J Virol 2016;90:5770-84.
- Goto H, Ohta K, Matsumoto Y. Evidence that receptor destruction by the Sendai virus hemagglutinin-neuraminidase protein is responsible for homologous interference. J Virol 2016;90:7640-6.
- Zeng Q, Langereis MA, van Vliet AL, Huizinga EG, de Groot RJ. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc Natl Acad Sci U S A 2008;105:9065-9.
- Hofling K, Klenk HD, Herrler G. Inactivation of inhibitors by the receptor-destroying enzyme of influenza C virus. J Gen Virol 1997;78:567-70.
- Kristiansen M, Froystad MK, Rishovd AL, Gjoen T. Characterization of the receptor-destroying enzyme activity from infectious salmon anaemia virus. J Gen Virol 2002;83:2693-7.
- Mei M, Ye J, Qin A, Wang L, Hu X, Qian K, et al. Identification of novel viral receptors with cell line expressing viral receptor-binding protein. Sci Rep 2015;5:7935.
- Kovacs D, Igaz N, Keskeny C, Belteky P, Toth T, Gaspar R, et al. Silver nanoparticles defeat p53-positive and p53-negative osteosarcoma cells by triggering mitochondrial stress and apoptosis. Sci Rep 2016;6:27902.
- Zielinska E, Zauszkiewicz-Pawlak A, Wojcik M, Inkielewicz-Stepniak I. Silver nanoparticles of different sizes induces a mixed type of programmed cell death in human pancreatic ductal adenocarcinoma. Oncotarget 2018;9:4675-97.
- Buttacavoli M, Albanese NN, Di Cara G, Alduina R, Faleri C, Gallo M, et al. Anticancer activity of biogenerated silver nanoparticles: An integrated proteomic investigation. Oncotarget 2018;9:9685-705.
- Ma DD, Yang WX. Engineered nanoparticles induce cell apoptosis: Potential for cancer therapy. Oncotarget 2016;7:40882-903.
- Shields DJ, Niessen S, Murphy EA, Mielgo A, Desgrosellier JS, Lau SK, et al. RBBP9: A tumor-associated serine hydrolase activity required for pancreatic neoplasia. Proc Natl Acad Sci U S A 2010;107:2189-94.
- Kirkegaard T, Jaattela M. Lysosomal involvement in cell death and cancer. Biochim Biophys Acta 2009;1793:746-54.
- Misaghi S, Ottosen S, Izrael-Tomasevic A, Arnott D, Lamkanfi M, Lee J, et al. Association of C-terminal ubiquitin hydrolase BRCA1-associated protein 1 with cell cycle regulator host cell factor 1. Mol Cell Biol 2009;29:2181-92.
- Kim HJ, Magesh V, Lee JJ, Kim S, Knaus UG, Lee KJ, et al. Ubiquitin C-terminal hydrolase-L1 increases cancer cell invasion by modulating hydrogen peroxide generated via NADPH oxidase 4. Oncotarget 2015;6:16287-303.
- Zhang F, Zhu X, Gong J, Sun Y, Chen D, Wang J, et al. Lysosome-mitochondria-mediated apoptosis specifically evoked in cancer cells induced by gold nanorods. Nanomedicine (Lond) 2016;11:1993-2006.
- Miyayama T, Matsuoka M. Involvement of lysosomal dysfunction in silver nanoparticle-induced cellular damage in A549 human lung alveolar epithelial cells. J Occup Med Toxicol 2016;11:1.
- Shen Y, Wu C, Uyeda TQP, Plaza GR, Liu B, Han Y, et al. Elongated nanoparticle aggregates in cancer cells for mechanical destruction with low frequency rotating magnetic field. Theranostics 2017;7:1735-48.
- EI-Ghonemy DE. Microbial amidases and their industrial applications: A review. J Med Microbiol Diagn 2014;4:1-6.
- Vago R, Bettiga A, Salonia A, Ciuffreda P, Ottria R. Development of new inhibitors for N-acylethanolamine-hydrolyzing acid amidase as promising tool against bladder cancer. Bioorg Med Chem 2017;25:1242-9.
- Melton RG, Sherwood RF. Antibody-enzyme conjugates for cancer therapy. J Natl Cancer Inst 1996;88:153-65.
- Romero P, Lopez R, Garcia E. Characterization of lytA-like N-acetylmuramoyl-L-alanine amidases from two new Streptococcus mitis bacteriophages provides insights into the properties of the major pneumococcal autolysin. J Bacteriol 2004;186:8229-39.
- Morales M, Martin-Galiano AJ, Domenech M, Garcia E. Insights into the evolutionary relationships of lytA autolysin and ply pneumolysin-like genes in Streptococcus pneumoniae and related streptococci. Genome Biol Evol 2015;7:2747-61.
- Sandalova T, Lee M, Henriques-Normark B, Hesek D, Mobashery S, Mellroth P, et al. The crystal structure of the major pneumococcal autolysin lytA in complex with a large peptidoglycan fragment reveals the pivotal role of glycans for lytic activity. Mol Microbiol 2016;101:954-67.
- Wang B, Chen Z, Yu F, Chen Q, Tian Y, Ma S, et al. Hsp90 regulates autophagy and plays a role in cancer therapy. Tumour Biol 2016;37:1-6.
- Singh PK, Donovan DM, Kumar A. Intravitreal injection of the chimeric phage endolysin ply187 protects mice from Staphylococcus aureus endophthalmitis. Antimicrob Agents Chemother 2014;58:4621-9.
- Guo M, Feng C, Ren J, Zhuang X. A novel antimicrobial endolysin, LysPA26, against Pseudomonas aeruginosa. Front Microbiol 2017;8:1-9.
- Gaupp R, Ledala N, Somerville GA. Staphylococcal response to oxidative stress. Front Cell Infect Microbiol 2012;2:33.
- Kashyap DR, Rompca A, Gaballa A, Helmann JD, Chan J, Chang CJ, et al. Peptidoglycan recognition proteins kill bacteria by inducing oxidative, thiol, and metal stress. PLoS Pathog 2014;10:e1004280.
- Wang AH, Hakoshima T, van der Marel G, van Boom JH, Rich A. AT base pairs are less stable than GC base pairs in Z-DNA: The crystal structure of d(m5CGTAm5CG). Cell 1984;37:321-31.